Without genetic variability, all living things should (by inheritance) be the same as the first. For having unequal beings, the only explanations would be those relating to single creations. But we know that the structure of DNA, which is at the basis of the transmission of hereditary characters, has a relative rather than absolute stability. While the stability guarantees the conservation of the initial information, the instability determines the modifications, or better (to use the specific term) the mutations.
Mutations can be divided into 3 large groups:
- gene mutations;
- chromosomal mutations;
- genomic mutations.
At this point it is good to briefly add two concepts: one is that of "repair" and the other is that of "favorable" or "unfavorable" mutation.
The repair, an English term that means repair, refers to the existence in the cells of specific enzymatic processes intended to verify the conservation of information, eliminating or correcting the stretches of DNA that do not conform to the original.
The concept of "favorable" or "unfavorable" mutation expresses the comparison between the efficiency, for the organism and for the species, of the original gene ("wild", ie "wild", primitive) and of the mutant gene.
It should be noted that advantage and disadvantage always refer to certain environmental conditions; a disadvantageous mutation in a certain environment can become favorable under different conditions.
The consequences of this principle are vast in population genetics, as can already be demonstrated by the different survival efficiencies of hemoglobin variants in normal or malarial environments. But this topic should be deferred to more detailed articles on general, human or medical genetics.
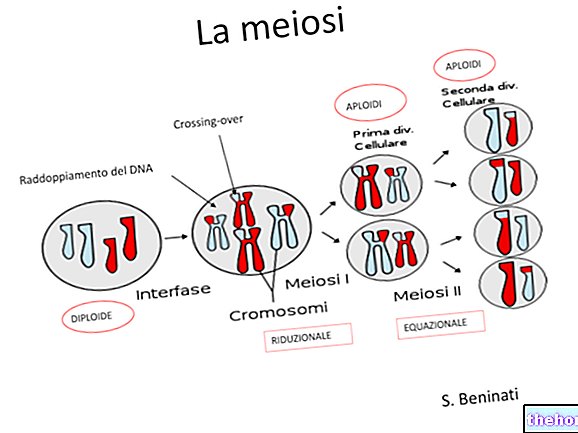
Another distinction is required (especially for gene mutations) between somatic mutations and germline mutations. All the cells of our multicellular organism derive, as is known, from a single zygote, but very early they differentiate into a somatic line, from which the whole body develops, and a germ line destined to form the gonads and, for terminal meiosis, the gametes It is quite evident that, except for interactions between the two cell populations, a mutation of the somatic line will manifest itself in the single organism but will not affect the descendants, while a mutation in the germline will only occur in the descendants.
Regarding the different types of mutation and the relative consequences, it is worth mentioning again a classification of the causes of mutation. These causes are called mutagens, which are used to distinguish mainly into physical and chemical mutagens. Various modifications of the physical environment can lead to mutations, but the main physical mutagens are radiation. For this reason radioactive substances are dangerous, and it is above all the heavier radioactive particles that tend to cause mutations by defection, with the most serious consequences .
Chemical mutagens can operate both by altering the ordered structure of nucleic acids, and by introducing into cells substances similar to normal nitrogenous bases, which can compete with the latter in the synthesis of nucleic acids, thus causing mutations by substitution.
GENE MUTATIONS
Gene mutations affect one or a few genes, that is, a limited stretch of DNA. Since information is stored in a sequence of nucleotide pairs, the smallest mutation unit (a muton) comprises a single complementary base pair. Without going into detail about the different mutation mechanisms at the gene level, we can limit ourselves to mentioning two: base substitution and reelection (or insertion). In base substitution mutations, one or more DNA nucleotides are replaced by others. If the error is not corrected in time, a sequence will follow at the time of transcription also altered in RNA. If the alteration of the triplet is not limited to a synonym (see the genetic code), it will result in the substitution of one or more amino acids in the polypeptide sequence. The substitution of an amino acid can be more or less critical for the determination of the structure of the protein and for its function.
In reelection or insertion mutations, one or more nucleotides are removed or added to the DNA sequence. These mutations are generally very serious because (unless they are whole triplets that add or subtract single amino acids) all the triplets that follow in the reading order are changed.
Gene mutations are the most frequent and are at the origin of most of the variability of hereditary traits between individuals.
CHROMOSOMIC MUTATIONS
These are mutations affecting relatively long fragments of a chromosome. It is customary to classify them mainly in:
- chromosomal mutations for re-election;
- chromosomal mutations by duplication;
- chromosomal mutations by translocation.
Mutations due to defection occur due to the breakage and loss of a more or less long fragment of chromosome. Especially in meiosis, this type of mutation is often lethal, due to the total loss of a certain number of genes, more or less indispensable.
In duplication mutations, following a break, the chromatid stumps tend to fuse.
In the subsequent removal of the centromeres, the chromosome, which has become dicentric, breaks into often unequal parts: as we can see, the result is re-election on the one hand and duplication on the other.
A chromosomal break can be followed by a reversal. The total genetic material is unchanged, but the sequence of the genes on the chromosome is altered.
The case of a translocation is similar, but it concerns the bonding of a fragment of a chromosome to a non-homologous chromosome. One chromosome is amputated and the other lengthened; the total genetic information of the cell is still unchanged, but the position effect is even more marked. It is easy to represent a position effect by referring to the concept of regulation of gene action: by changing position on chromosomes, a gene can easily leave one operon and be inserted into another, resulting in an "altered activation or repression.
However, it is said that a translocation is balanced (or balanced) when there is reciprocity of translocation between two pairs of chromosomes, keeping the sum of the gene information unchanged. The cross-shaped figure in meiotic diakinesis generally corresponds to the balanced translocation.
GENOMIC MUTATIONS
Remembering that the genome is the individual genetic patrimony, ordered in chromosomes, it can be specified that we speak of genomic mutations when the chromosomes have a different distribution from the norm of the species.
Genomic mutations can be classified mainly in mutations for polyploidy, haploidy and aneuploidy.
Polyploidy mutations occur when reduplication does not result in division; they occur more easily in plants, where indeed they are used to improve production.
If cell division fails in meiosis, diploid gametes may occur; if such a gamete manages to merge with a normal gamete, the zygote resulting from this fertilization will be triploid. Such a zygote can sometimes succeed in originating a whole organism, since reduplication and mitosis do not require an even number of chromosomes. At the time of meiosis, however, regular pairing of homologous chromosomes will be impossible.
Genomic mutations by haploidy can occur when, in a normally diploid species, a gamete is activated by another gamete devoid of nuclear material, or even in the absence of fertilization: a haploid individual will result.
While the previous types of genomic mutations always concern the addition or subtraction of whole numbers n of chromosomes, mutations for aneuploidy concern the excess or defect of single chromosomes (chromosomal aberrations).
The euploid chromosomal makeup of a species is defined as its normal karyotype or idiotype.