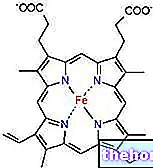
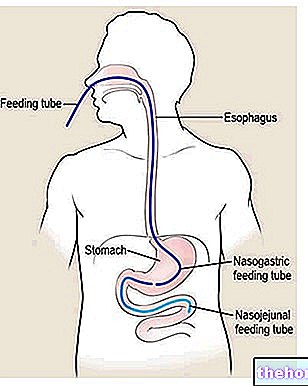
The krebs cycle is also called the tricarboxylic acid cycle and uses acetyl coezime A as its starting metabolite, which is obtained by the action of pyruvate dehydrogenase on the pyruvate produced by glycolysis.
ATP and reducing power are obtained from the krebs cycle; the reducing power is sent to the respiratory chain where NADH and FADH2 are oxidized respectively to NAD + and FAD: the reducing power is transferred, along the respiratory chain, to coupling systems from which further ATP is produced.
The krebs cycle is a nerve center not only for glucose metabolism but also for the metabolism of fatty acids and amino acids, in fact the pyruvate that is converted to acetyl coenzyme A does not come only from the degradation of glucose: it is obtained, for example , also from the transamination of alanine (an amino acid).
About 80% of the acetyl coenzyme A that participates in the krebs cycle comes from the metabolism of fatty acids.
Acetyl coenzyme A is a thioester therefore it has a high energy content which is exploited by citrate synthase to form a new carbon-carbon bond; citrate synthase is the first enzyme of the krebs cycle.
The methyl carbon of acetyl coenzyme A willingly releases (by tautomerism) a proton (becomes a carboanion) and attacks the carbonyl carbon of oxaloacetate: a high-energy thioester (citril coenzyme A) is formed from which, by hydrolysis, the citrate is obtained and coenzyme A is reformed. The citrate synthase is negatively modulated by the product, that is by the citrate and by the ATP: if the citrate accumulates it means that this stage is faster than the others therefore it must slow down (citrate is a negative modulator).

ATP also influences the action of citrate synthase since the reducing power is obtained from the krebs cycle which is then sent to the respiratory chain from which ATP is produced; if ATP accumulates it means that more than what is needed is produced. By slowing down the krebs cycle (the cycle slows down if one of its stages is slowed down) the production of ATP is also slowed down: the negative modulation of ATP is a feed-back modulation (the formation of one of the final products is modulated by adjusting the speed of one step in the process).
In the second stage of the krebs cycle, the citrate is converted into isocitrate by the action of the enzyme aconitase; the name of the enzyme derives from the fact that the citrate is first dehydrated with the formation of cis-aconitate and, subsequently, the water re-enters by attaching itself to a carbon different from the one on which it was previously bonded. The isocitrate is obtained without the substrate leaving the catalytic site; aconitase is a stereospecific enzyme: it recognizes the three carboxyl centers of the citrate and this causes the citrate to remain bound to the enzyme so that the exit and entry of the " water always passes through the cis-aconitate intermediate.
In the third stage of the krebs cycle there is the first energy consideration because there is the loss of a carbon eliminated as carbon dioxide. The enzyme that catalyzes this stage is the isocitrate dehydrogenase; the substrate undergoes, first of all, a dehydrogenation: the NAD + acquires reducing power and oxalosuccinate is formed (it is an oxal derivative of succinic acid). The oxalosuccinate then undergoes decarboxylation to α-ketoglutarate.
The isocitrate dehydrogenase enzyme has two modulation sites: a positive modulation due to ADP and a negative modulation due to ATP. The amount of ATP consumed daily is very high: ATP provides the energy released by its hydrolysis, "ADP and all" orthophosphate.
The total concentration of nucleosides (nitrogenous base plus sugar) and nucleotides (nucloside plus phosphate) in an organism is almost constant: to say, therefore, that c "is a lot of ATP or little ADP (or vice versa, a lot of ADP and little ATP) is the same thing; ADP is a synonym for energy need and is therefore a positive modulator, while ATP is a symptom of energy availability and is therefore a negative modulator.
CONTINUE: Second part "